Evolución, Embriología y Estructura del Sistema Nervioso
1. Evolución del Sistema Nervioso
Los primeros animales unicelulares o protozoarios tuvieron irritabilidad primitiva y cuando la evolución produjo los primeros organismos pluricelulares o metazoarios aparecieron por primera vez células adaptadas específicamente para la interacción intercelular. Estas células fueron las causantes de la irritabilidad en los organismos. Ahora la principal función del sistema nervioso era la coordinación de estímulos y diseminar respuestas simples. Pero cuando evolucionó el sistema nervioso de los vertebrados, surgió de una red de comunicación simple, el Sistema Nervioso Periférico, que se extendía a todas las partes del cuerpo y un aparato de coordinación, el Sistema Nervioso Central. Los gusanos desarrollaron un sistema nervioso semejante a un cordón que corre a lo largo de su cuerpo con un gran centro nervioso o cerebro que contiene neuronas de asociación en su extremidad anterior. Indudablemente este fue el precursor del cerebro de los vertebrados y de la médula espinal. El primer rasgo de la evolución fue la aparición de nervios aferentes y eferentes en el sistema nervioso periférico. A medida que los animales fueron evolucionando adquirieron receptores a distancia: ojos, olfato y oído en las extremidades anteriores de sus cuerpos. El cerebro apareció muy temprano como un pequeño abultamiento, sin embargo esta fue la porción del encéfalo que más se desarrolló en la evolución. La mayor complejidad fue el hecho de anteponer nuevas estructuras sobre las antiguas, por ejemplo, se añadió el tronco encefálico a la médula espinal y el cerebelo al tronco encefálico. La evolución del sistema nervioso añadió flexibilidad y adaptabilidad al estímulo-respuesta a medida que crecían las presiones ambientales.
Invertebrados
Animales sin sistema nervioso son capaces de tener conductas realmente complejas.
Vertebrados
En estos la médula espinal no evolucionó tanto, pero en todos los vertebrados es el centro del comportamiento reflejo y conduce impulsos entre los órganos sensoriales y músculos por una parte y por otra el cerebro. Pero esto no quiere decir que no haya tenido cambios, por ejemplo, los nervios espinales regulan tanto en peces inferiores y hombre, pero en aquellos les corresponde mediar también funciones autónomas y solo en peces complejos hay ganglios simpáticos junto con sistema autónomo bien diferenciado.
2. Embriología
Segmentación: 2-16 células
Mórula: 16 a 60 células
Se dividen dos grupos de células:
- Trofoblastos: que son las que dan origen a la placenta que es la que se encarga de la nutrición.
Blástula:
Las células se dividen en tres grupos:
- Ectodermo: Sistema nervioso
- Mesodermo: Sistema urinario
- Endodermo: Tubo digestivo, hígado, páncreas
Se invaginará:
Tubo neural y crestas neurales:
Del tubo se generará el Sistema Nervioso Central constituido por encéfalo y médula espinal.
Crestas:
- Región dorsal: aferentes o sensitivas
- Región ventral: eferentes o motoras
Del tubo neural se producen tres vesículas primarias y de estas salen 5 secundarias:
- Prosencéfalo:
- Telencéfalo: Hemisferios cerebrales
- Diencéfalo: Tálamo e Hipotálamo
- Mesencéfalo: Pedúnculos cerebrales, lámina cuadrigémina
- Rombencéfalo:
- Metencéfalo: Puente y cerebelo
- Mielencéfalo: Médula oblonga o bulbo raquídeo
3. La Neurona
Las neuronas son un tipo de células del sistema nervioso cuya principal característica es la excitabilidad de su membrana plasmática; están especializadas en la recepción de estímulos y conducción del impulso nervioso (en forma de potencial de acción) entre ellas o con otros tipos celulares, como por ejemplo las fibras musculares de la placa motora. Altamente diferenciadas, la mayoría de las neuronas no se dividen una vez alcanzada su madurez; no obstante, una minoría sí lo hace. Las neuronas presentan unas características morfológicas típicas que sustentan sus funciones: un cuerpo celular o «pericarion», central; una o varias prolongaciones cortas que generalmente transmiten impulsos hacia el soma celular, denominadas dendritas; y una prolongación larga, denominada axón o «cilindroeje», que conduce los impulsos desde el soma hacia otra neurona u órgano diana.
La neurogénesis en seres adultos, fue descubierta apenas en el último tercio del siglo XX. Hasta hace pocas décadas se creía que, a diferencia de la mayoría de las otras células del organismo, las neuronas normales en el individuo maduro no se regeneraban, excepto las células olfatorias. Los nervios mielinados del sistema nervioso periférico también tienen la posibilidad de regenerarse a través de la utilización del neurolema, una capa formada de los núcleos de las células de Schwann.
Núcleo
Situado en el cuerpo celular, suele ocupar una posición central y ser muy conspicuo (visible), especialmente en las neuronas pequeñas. Contiene uno o dos nucléolos prominentes, así como una cromatina dispersa, lo que da idea de la relativamente alta actividad transcripcional de este tipo celular. La envoltura nuclear, con multitud de poros nucleares, posee una lámina nuclear muy desarrollada. Entre ambos puede aparecer el cuerpo accesorio de Cajal, una estructura esférica de en torno a 1 μm de diámetro que corresponde a una acumulación de proteínas ricas en los aminoácidos arginina y tirosina.
Pericarion
Diversos organelos llenan el citoplasma que rodea al núcleo. El organelo más notable, por estar el pericarion lleno de ribosomas libres y adheridos al retículo rugoso, es la llamada sustancia de Nissl, al microscopio óptico, se observan como grumos basófilos, y, al electrónico, como apilamientos de cisternas del retículo endoplasmático. Tal abundancia de los orgánulos relacionados en la síntesis proteica se debe a la alta tasa biosintética del pericarion.
Estos son particularmente notables en neuronas motoras somáticas, como las del ucerno anterior de la médula espinal o en ciertos núcleos de nervios craneales motores. Los cuerpos de Nissl no solamente se hallan en el pericarion sino también en las dendritas, aunque no en el axón, y es lo que permite diferenciar de dendritas y axones en el neurópilo.
El aparato de Golgi, que se descubrió originalmente en las neuronas, es un sistema muy desarrollado de vesículas aplanadas y agranulares pequeñas. Es la región donde los productos de la sustancia de Nissl posibilitan una síntesis adicional. Hay lisosomas primarios y secundarios (estos últimos, ricos en lipofuscina, pueden marginar al núcleo en individuos de edad avanzada debido a su gran aumento). Las mitocondrias, pequeñas y redondeadas, poseen habitualmente crestas longitudinales.
En cuanto al citoesqueleto, el pericarion es rico en microtúbulos (clásicamente, de hecho, denominados neurotúbulos, si bien son idénticos a los microtúbulos de células no neuronales) y filamentos intermedios (denominados neurofilamentos por la razón antes mencionada). Los neurotúbulos se relacionan con el transporte rápido de las moléculas de proteínas que se sintetizan en el cuerpo celular y que se llevan a través de las dendritas y el axón.
Dendritas
Las dendritas son ramificaciones que proceden del soma neuronal que consisten en proyecciones citoplasmáticas envueltas por una membrana plasmática sin envuelta de mielina. En ocasiones, poseen un contorno irregular, desarrollando espinas. Sus orgánulos y componentes característicos son: muchos microtúbulos y pocos neurofilamentos, ambos dispuestos en haces paralelos; muchas mitocondrias; grumos de Nissl, más abundantes en la zona adyacente al soma; retículo endoplasmático liso, especialmente en forma de vesículas relacionadas con la sinapsis.
Axón
El axón es una prolongación del soma neuronal recubierta por una o más células de Schwann en el sistema nervioso periférico de vertebrados, con producción o no de mielina. Puede dividirse, de forma centrífuga al pericarion, en: cono axónico, segmento inicial, resto del axón.
- Cono axónico: Adyacente al pericarion, es muy visible en las neuronas de gran tamaño. En él se observa la progresiva desaparición de los grumos de Nissl y la abundancia de microtúbulos y neurfilamentos que, en esta zona, se organizan en haces paralelos que se proyectarán a lo largo del axón.
- Segmento inicial: En él comienza, de existir, la mielinización externa. En el citoplasma, a esa altura se detecta una zona rica en material electronodenso en continuidad con la membrana plasmática, constituido por material filamentoso y partículas densas; se asume que interviene en la generación del potencial de acción que transmitirá la señal sináptica. En cuanto al citoesqueleto, posee esta zona la organización propia del resto del axón. Los microtúbulos, ya polarizados, poseen la proteína τ pero no la proteína MAP-2.
- Resto del axón: En esta sección comienzan a aparecer los nódulos de Ranvier y las sinapsis.
Clasificación
Aunque el tamaño del cuerpo celular puede ser desde 5 hasta 135 micrómetros, las prolongaciones o dendritas pueden extenderse a una distancia de más de un metro. El número, la longitud y la forma de ramificación de las dendritas brindan un método morfológico para la clasificación de las neuronas.
Según el tamaño de las prolongaciones:
- Poliédricas: como las motoneuronas del asta anterior de la médula.
- Fusiformes: como las células de doble ramillete de la corteza cerebral.
- Estrelladas: como las neuronas aracniformes y estrelladas de la corteza cerebral y las estrelladas, en cesta y Golgi del cerebelo.
- Esféricas: en ganglios raquídeos, simpáticos y parasimpáticos
- Piramidales: presentes en la corteza cerebral.
Según la polaridad:
- Neuronas monopolares o unipolares: son aquéllas desde las que nace sólo una prolongación que se bifurca y se comporta funcionalmente como un axón salvo en sus extremos ramificados en que la rama periférica reciben señales y funcionan como dendritas y transmiten el impulso sin que este pase por el soma neuronal. Son típicas de los ganglios de invertebrados y de la retina.
- Neuronas bipolares: poseen un cuerpo celular alargado y de un extremo parte una dendrita y del otro el axón (solo puede haber uno por neurona). El núcleo de este tipo de neurona se encuentra ubicado en el centro de ésta, por lo que puede enviar señales hacia ambos polos de la misma. Ejemplos de estas neuronas se hallan en las células bipolares de la retina (conos y bastones), del ganglio coclear y vestibular, estos ganglios son especializados de la recepción de las ondas auditivas y del equilibrio.
- Neuronas multipolares: tienen una gran cantidad de dendritas que nacen del cuerpo celular. Ese tipo de células son la clásica neurona con prolongaciones pequeñas (dendritas) y una prolongación larga o axón. Representan la mayoría de las neuronas. Dentro de las multipolares, distinguimos entre las que son de tipo Golgi I, de axón largo, y las de tipo Golgi II, que no tienen axón o éste es muy corto. Las neuronas de proyección son del primer tipo, y las neuronas locales o interneuronas del segundo.
- Neuronas pseudounipolares: son aquéllas en las cuales el cuerpo celular tiene una sola dendrita o neurita, que se divide a corta distancia del cuerpo celular en dos ramas, motivo por cual también se les denomina pseudounipolares (pseudos en griego significa «falso»), una que se dirige hacia una estructura periférica y otra que ingresa en el sistema nervioso central. Se hallan ejemplos de esta forma de neurona en el ganglio de la raíz posterior.
- Neuronas anaxónicas: son pequeñas. No se distinguen las dendritas de los axones. Se encuentran en el cerebro y órganos especiales de los sentidos.
Según las características de las neuritas:
- De acuerdo a la naturaleza del axón y de las dendritas, clasificamos a las neuronas en:
- Axón muy largo o Golgi de tipo I. El axón se ramifica lejos del pericarion. Con axones de hasta 1 m.
- Axón corto o Golgi de tipo II. El axón se ramifica junto al soma celular.
- Sin axón definido. Como las células amacrinas de la retina.
- Isodendríticas. Con dendritas rectilíneas que se ramifican de modo que las ramas hijas son más largas que las madres.
- Idiodendríticas. Con las dendritas organizadas dependiendo del tipo neuronal; por ejemplo, como las células de Purkinje del cerebelo.
- Alodendríticas. Intermedias entre los dos tipos anteriores.
Según el mediador químico:
- Colinérgicas. Liberan acetilcolina.
- Noradrenérgicas. Liberan norepinefrina.
- Dopaminérgicas. Liberan dopamina.
- Serotoninérgicas. Liberan serotonina.
- GABAérgicas. Liberan GABA, es decir, ácido γ-aminobutírico.
4. La Sinapsis
La sinapsis (del gr. σύναψις, «enlace») es el proceso de comunicación entre neuronas. Se inicia con una descarga química que origina una corriente eléctrica en la membrana de la célula presináptica (célula emisora); una vez que este impulso nervioso alcanza el extremo del axón, la propia neurona segrega un tipo de proteínas (neurotransmisores) que se depositan en el espacio sináptico, espacio intermedio entre esta neurona transmisora y la neurona postsináptica (receptora). Estos neurotransmisores son los encargados de excitar o inhibir la acción de la otra neurona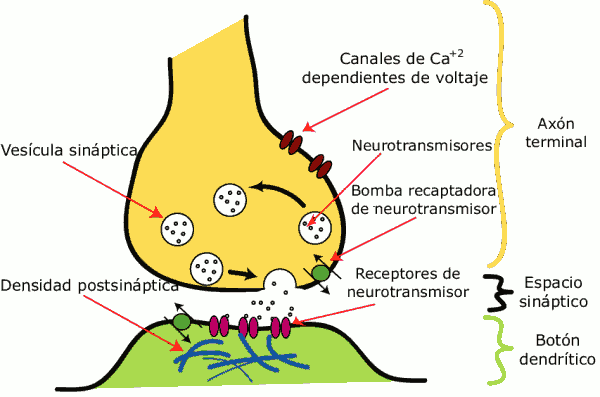
5. Neurotransmisores
Un neurotransmisor es una biomolécula, sintetizada generalmente por las neuronas, que se vierte, a partir de vesículas existentes en la neurona presináptica, hacia la brecha sináptica y produce un cambio en el potencial de acción de la neurona postsináptica. Los neurotransmisores son, por tanto, las principales sustancias de las sinapsis.
6. Potencial de Acción
Un potencial de acción o también llamado impulso eléctrico, es una onda de descarga eléctrica que viaja a lo largo de la membrana celular. Los potenciales de acción se utilizan en el cuerpo para llevar información entre unos tejidos y otros, lo que hace que sean una característica microscópica esencial para la vida de los animales. Pueden generarse por diversos tipos de células corporales, pero las más activas en su uso son las células del sistema nervioso para enviar mensajes entre células nerviosas o desde células nerviosas a otros tejidos corporales, como el músculo o las glándulas.
Muchas plantas también generan potenciales de acción que viajan a través del floema para coordinar su actividad. La principal diferencia entre los potenciales de acción de animales y plantas es que las plantas utilizan flujos de potasio y calcio mientras que los animales utilizan potasio y sodio.
Los potenciales de acción son la vía fundamental de transmisión de códigos neurales. Sus propiedades pueden frenar el tamaño de cuerpos en desarrollo y permitir el control y coordinación centralizados de órganos y tejidos.
Umbral e Iniciación
Los potenciales de acción se desencadenan cuando una despolarización inicial alcanza un umbral. Este potencial umbral varía, pero normalmente está en torno a -55 a -30 milivoltios sobre el potencial de reposo de la célula, lo que implica que la corriente de entrada de iones sodio supera la corriente de salida de iones potasio. El flujo neto de carga positiva que acompaña los iones sodio despolariza el potencial de membrana, desembocando en una apertura de los canales de sodio dependientes de voltaje. Estos canales aportan un flujo mayor de corrientes iónicas hacia el interior, aumentando la despolarización en una retroalimentación positiva que hace que la membrana llegue a niveles de despolarización elevados.
El umbral del potencial de acción puede variar cambiando el equilibrio entre las corrientes de sodio y potasio. Por ejemplo, si algunos de los canales de sodio están inactivos, determinado nivel de despolarización abrirá menos canales de sodio, y aumenta así el umbral de despolarización necesario para iniciar el potencial de acción. Esta es el principio del funcionamiento del periodo refractario.
Los potenciales de acción son muy dependientes de los equilibrios entre iones sodio y potasio (aunque hay otros iones que contribuyen minoritariamente a los potenciales, como calcio y cloro), y por ello los modelos se hacen utilizando sólo dos canales iónicos transmembrana: un canal de sodio dependiente de voltaje y un canal de potasio pasivo. El origen del umbral del potencial de acción puede visualizarse en la curva I/V que representa las corrientes iónicas a través de los canales frente al potencial de membrana.
El umbral del potencial de acción se confunde a veces con el umbral de la apertura de canales de sodio. Es una incorrección, ya que los canales de sodio carecen de umbral. Por el contrario, se abren en respuesta a la despolarización aleatoriamente. La despolarización no implica tanto la apertura de los canales como el incremento de la probabilidad de que se abran. Incluso en potenciales de hiperpolarización, un canal de sodio puede abrirse esporádicamente. Además, el umbral del potencial de acción no es el voltaje a la que el flujo de iones sodio se hace importante; es el punto en que excede el flujo de potasio.
Biológicamente, en las neuronas la despolarización se origina en las sinapsis dendríticas. En principio, los potenciales de acción podrían generarse en cualquier punto a lo largo de la fibra nerviosa. Cuando Luigi Galvani descubrió la electricidad animal haciendo que la pierna de una rana muerta volviese a la vida tocando el nervio ciático con un escalpelo, aplicándole sin darse cuenta una carga electrostática negativa e iniciando un potencial de acción.
7. Microglía
Las microglías son células pequeñas con núcleo alargado y con prolongaciones cortas e irregulares que tienen capacidad fagocitaria. Se originan en precursores de la médula ósea y alcanzan el sistema nervioso a través de la sangre; representan el sistema mononuclear fagocítico en el sistema nervioso central.
Contienen lisosomas y cuerpos residuales. Generalmente se la clasifica como célula de la neuroglia. Presentan el antígeno común leucocítico y el antígeno de histocompatibilidad clase II, propio de las células presentadoras de antígeno.